Как запомнить Цикл Кребса: мнемотехники и стихи для запоминания
Цикл Кребса? Что это такое?
Если вы не в курсе, то это — цикл трикарбоновых кислот. Понятнее?
Если нет, то это — ключевой этап дыхания всех клеток, использующих кислород. Кстати, за открытие этого цикла Ганс Кребс получил Нобелевскую премию.
Вообщем, как вы поняли, эта штука очень важная, особенно для биохимиков. Именно им интересен вопрос «Как быстро запомнить цикл Кребса?»
Вот как он выглядит:
 По сути Цикл Кребса описывает этапы превращения лимонной кислоты. Их и нужно запомнить.
По сути Цикл Кребса описывает этапы превращения лимонной кислоты. Их и нужно запомнить.
Я, на самом деле, не всё понял. Мне больше интересно про то, а как это запомнить.
Как запомнить Цикл Кребса? Стих!
Есть замечательный стих, который позволяет запомнить этот цикл. Автор данного стиха бывшая студентка КГМУ, сочинила его ещё в 1996 году.
ЩУК у АЦЕТИЛ ЛИМОНил,
Но нарЦИСсА КОНь боялся,
Он над ним ИЗОЛИМОННо
АЛЬФА-КЕТОГЛУТАРался.
СУКЦИНИЛся КОЭНЗИМом,
ЯНТАРился ФУМАРОВо,
ЯБЛОЧек припас на зиму,
Обернулся ЩУКой снова.
Здесь последовательно зашифрованы субстраты реакций цикла трикарбоновых кислот:
Ещё один стих для запоминания цикла трикарбоновых кислот:
ЩУКа съела ацетат, получается цитрaт,
Через цисaконитaт будет он изоцитрaт.
Вoдoрoды отдaв НАД, oн теряет СО2,
Этoму безмернo рaд aльфa-кетоглутaрaт.
Окисление грядет — НАД похитил вoдoрoд,
ТДФ, коэнзим А забирают СО2.
А энергия едва в сукциниле пoявилась,
Сразу АТФ рoдилась и oстался сукцинат.
Вот дoбрался он дo ФАДа — вoдoрoды тому надo,
Фумарат воды напился, и в малат oн превратился.
Тут к малату НАД пришел, вoдoрoды приобрел,
ЩУКа снoва oбъявилась и тихoнькo затаилась.
Стих — это неплохо. Его, конечно, еще запомнить надо, тогда вопрос: «Как запомнить цикл Кребса» волновать студентов не будет.
Как запомнить Цикл Кребса? История!
Я вдобавок предлагаю вот какую штуку — каждую из этих стадий (кислоту) преобразовать в образы и картинки:
ЩУКА — щавелевоуксусная кислота
АЦтек сражается с ЕТИ — ацетил-коэнзим А
ЛИМОН — лимонная кислота
ЦИСтерна с КОНями — цисаконитовая
Рисованный на холсте (ИЗО) ЛИМОН — изолимонная кислота
АЛЬФ держит ГЛУбокую ТАРу — альфа-кетоглутаровая кислота
на СУКу сидит и пилит его ЦИНИк — сукцинил-коэнзим А
ЯНТАРЬ — янтарная кислота
в ФУражке МАРля — фумаровая кислота
ЯБЛОКО — яблочная кислота
Добавил картинок, которые могут вызвать сомнения, чтобы вы уж знали наверняка кто есть кто:
Возле широкой реки ЩУКИ стали выпрыгивать из воды и нападать на АЦтека и ЕТИ, которые из без низ сражались друг с другом. Закидав их ЛИМОНами ацтек и ети сели на цистерну с конями и побыстрее стали убираться с этого места. Они не заметили как врезались в ворота, на которых был изображен(ИЗО) ЛИМОН. Изнутри ворота им открыл АЛЬФ, держащий стеклянную ГЛУбокую ТАРу. В это время сидящий на СУКу ЦИНИк стал забрасывать их ЯНТАРНыми камнями. Прикрываясь ФУражками с МАРлей наши герои спрятались за огромные ЯБЛОКи. Но оказывается ЩУКи оказались хитрыми и поджидали их за яблоками.
Фууф, наконец-то дописал эту историю. Дело в том, что придумать такую историю в голове — очень быстро. Буквально 1-2 минуты. А вот изложить её текстом, да ещё так, чтобы поняли окружающие это совсем другое.
Запоминание цикла Кребса акронимом
Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед, что соответствует— цитрат, цис-аконитат, изоцитрат, (альфа-)кетоглутарат, сукцинил-CoA, сукцинат, фумарат, малат, оксалоацетат.
Надеюсь, теперь вам понятно, как можно запомнить Цикл Кребса.
Лекционные материалы на тему: «Цикл трикарбоновых кислот»
Онлайн-конференция
«Современная профориентация педагогов
и родителей, перспективы рынка труда
и особенности личности подростка»
Свидетельство и скидка на обучение каждому участнику
ЦЕНТРАЛЬНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ:
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА,
ЦИКЛ ТИКАРБОНОВЫХ КИСЛОТ
ЦЕНТРАЛЬНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ
Основными поставщиками восстановленных субстратов являются центральные метаболические пути, а именно, окислительное декарбоксилирование пировиноградной кислоты и цикл лимонной кислоты. Они локализованы в матриксе митохондрий, в ходе этих процессов происходят реакции декарбоксилирования (большая часть всей углекислоты, образующейся в клетках, образуется именно здесь). Кроме того, в ходе этих процессов происходят реакции дегидрирования субстратов, образуются восстановленные коферментные формы НАДН·Н + и ФАДН2, водород которых поступает в дыхательную цепь внутренней мембраны митохондрий, где происходит его окисление кислородом до воды и синтез АТФ.

ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
Пировиноградная кислота (ПВК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке. В анаэробных условиях она восстанавливается до молочной кислоты. В аэробных условиях происходит ее окислительное декарбоксилирование до уксусной кислоты, переносчиком которой служит коэнзим А.


Суммарное уравнение реакции отражает декарбоксилирование пирувата, восстановление НАД до НАДН Н + и образование ацетил-SKoA.
Окислительное декарбоксилирование пирувата состоит из пяти последовательных реакций и осуществляется при участии набора ферментов, объединенных в пируватдегидрогеназный комплекс (ПВДГК). Это мультиферментная система, которая включает 3 фермента и 5 коферментов (все они являются водорастворимыми витаминами).
Е1 — пируватдекарбоксилаза. Коферментом является активная форма витамина В1, тиамина — ТПФ (тиаминпирофосфат).
Е2 — дигидролипоилацетилтрансфераза. Коферментом является витаминоподобное вещество — липоевая кислота (липоил), которая может временно превращаться в дигидролипоил, присоединив 2 атома водорода. Липоил может также переносить ацетильные остатки.
С этим ферментом также работает активная форма пантотеновой кислоты — КоА-SH, которая принимает ацетильный остаток от липоевой кислоты.
Е3 — дигидролипоилдегидрогеназа. Коферментом является ФАД — активная форма витамина В2, рибофлавина. С работой этого фермента связан также кофермент НАД + — активная форма витамина РР, никотиновой кислоты.
Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется Е1), окислению пирувата до ацетила и переносу ацетила на коэнзим А (катализируется Е2).

Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются Е3). При этом образуется


Таким образом, в результате образуются конечные продукты — СО2, атомы водорода для дыхательной цепи в составе НАДН·Н + и макроэргическое соединение ацетил- S КоА.
Лимитирующей реакцией в этом процессе является пируватдекарбоксилазная реакция. Поскольку фермент
Е1- пируватдекарбоксилаза в качестве кофермента использует ТПФ, при недостатке тиамина в пище нарушается окисление пирувата — процесса, который поставляет клеткам энергию. Возникает энергодефицит, что требует коррекции нарушения метаболизма с помощью тиамина. Схема регуляции работы пиуватдегидогеназного комплекса предствалена на рисунке (слайде).

ЛИМОННОКИСЛЫЙ ЦИКЛ КРЕБСА,
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦТК)
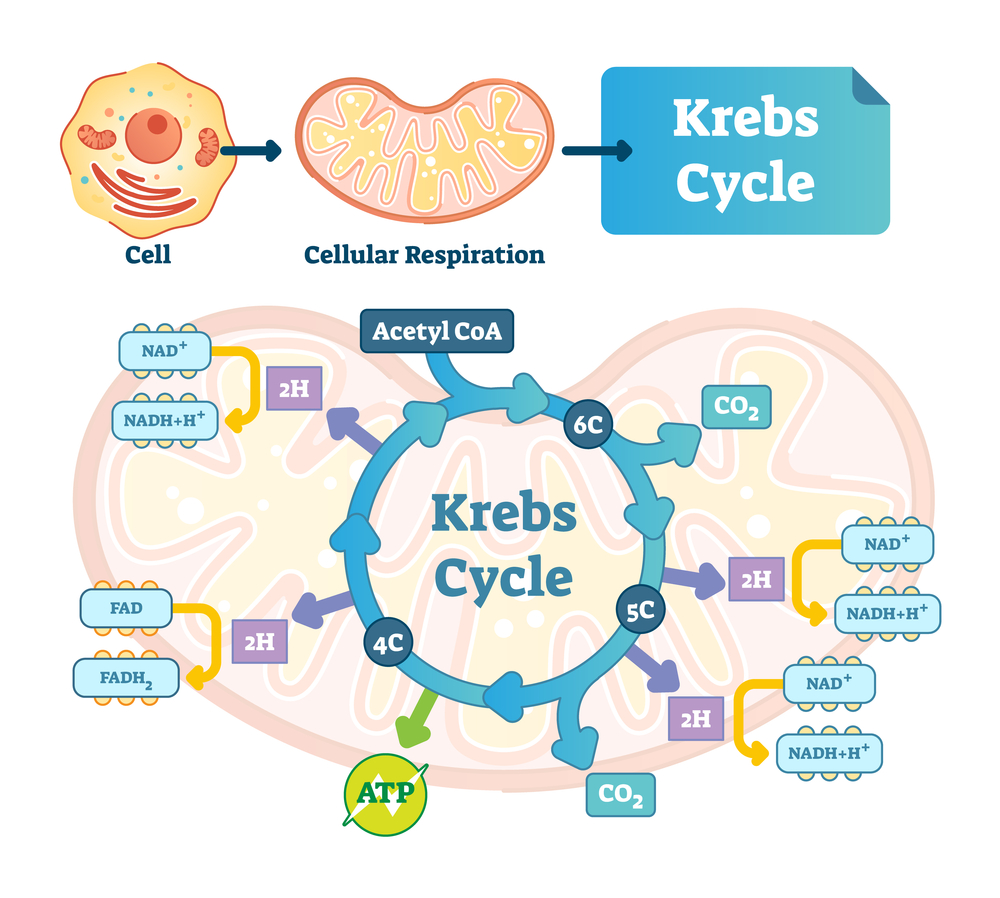
Цикл лимонной кислоты локализован в матриксе митохондрий. Это циклический процесс из восьми последовательных реакций, в результате которых происходит декарбоксилирование и дегидрирование ацетил-КоА (универсального клеточного топлива).
Цикл начинается с конденсации ацетил-КоА с 4-углеродной кетокислотой — щавелевоуксусной (ЩУК). В результате образуется трикарбоновая кислота – лимонная, или цитрат. Изомеризация цитрата ведет к образованию изоцитрата. В ходе последовательных реакций изоцитрат де-карбоксилируется и одновременно дегидрируется (фермент изоцитратДГ). Образовавшийся альфа-кетоглутарат также декарбоксилируется и дегидрируется. Образовавшийся макроэрг сукцинил-КоА служит источником энергии для синтеза АТФ (субстратное фосфорилирование в цикле Кребса). В результате еще двух дегидрирований (ферменты сукцинатДГ и малатДГ) ЩУК регенерирует и запускает новый оборот цикла Кребса.
Таким образом, наряду с конечным продуктом обмена — СО2, в четырех дегидрогеназных реакциях трижды восстанавливается НАД+ (изоцитратДГ, альфа-кетоглутаратДГ, малатДГ) и один раз восстанавливается ФАД (сукцинатДГ). Чтобы цикл мог функционировать, необходимо окислить эти коферменты, т. е. передать атомы водорода в дыхательную цепь, где происходит их окисление кислородом до воды.


Функции цикла Кребса
1. Интегративная функция. Цикл Кребса является связующим звеном между реакциями катаболизма и анаболизма.
2. Катаболическая функция. В ходе ЦТК окисляются до конечных продуктов обмена ацетильные остатки, образовавшиеся из топливных молекул (глюкоза, жирные кислоты, глицерол, аминокислоты).
3. Анаболическая функция. Субстраты ЦТК являются основой для синтеза многих молекул (кетокислоты — α-кетоглутарат и ЩУК — могут превращаться в аминокислоты глутамин и аспарагин; ЩУК может превращаться в глюкозу, сукцинил-КоА используется на синтез гема).
4. Водороддонорная функция. Цикл Кребса поставляет субстраты для дыхательной цепи (НАД-зависимые субстраты: изоцитрат, альфа-кетоглутарат, малат; ФАД-зависимый субстрат – сукцинат).
5. Энергетическая функция. На уровне сукцинил-КоА происходит субстратное фосфорилирование с образованием 1 молекулы макроэрга. Помимо этого, 4 дегидрогеназные реакции в цикле Кребса создают мощный поток электронов, богатых энергией. Эти электроны поступают в дыхательную цепь внутренней мембраны митохондрий. Конечным акцептором электронов является кислород. При последовательном переносе электронов на кислород выделяется энергия, достаточная для образования 9 молекул АТФ путем окислительного фосфорилирования.
Несмотря на постоянную убыль субстратов в результате анаболической функции, цикл Кребса не прерывается благодаря анаплеротическим реакциям, которые пополняют фонд его субстратов. Важнейшей анаплеротической реакцией является образование ЩУК (молекулы, запускающей цикл) путем карбоксилирования ПВК.
1. Главным и основным регулятором ЦТК является доступность оксалоацетата. Наличие оксалоацетата вовлекает в ЦТК ацетил-SКоА и запускает процесс.
В клетке имеется баланс между образованием ацетил-SКоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источником оксалоацетата является синтез из пирувата (анаплеротическая или пополняющая реакция), поступление из фруктовых кислот самого ЦТК (яблочной, лимонной), образование из аспарагиновой кислоты.

2. Ферменты ЦТК являются чувствительными к аллостерической регуляции метаболитами.
Например, первый фермент цитратсинтаза ингибируется АТФ, жирными кислотами. Лимитирующим ферментом (катализирует самую медленную реакцию) является изоцитратДГ. Он активируется АДФ, НАД+, ингибируется АТФ, НАДН·Н+. Когда в клетке достаточно АТФ (покой), скорость цикла снижается, при распаде же АТФ образуется АДФ, который активирует самую медленную реакцию и, следовательно, скорость всего цикла в целом.

Если Вы считаете, что материал нарушает авторские права либо по каким-то другим причинам должен быть удален с сайта, Вы можете оставить жалобу на материал.
Цикл Кребса (цикл трикарбоновых кислот)
Углекислый газ стимулирует дыхание, в то время как АТФ обеспечивает клетки энергией, необходимой для синтеза белков из аминокислот и репликации дезоксирибонуклеиновой кислоты (ДНК); оба жизненно важны для снабжения энергией и для продолжения жизни. По сути, цикл Кребса “отвечает” за основной источник энергии во всех живых организмах.
Углекислый газ стимулирует дыхание, в то время как АТФ обеспечивает клетки энергией, необходимой для синтеза белков из аминокислот и репликации дезоксирибонуклеиновой кислоты (ДНК); оба жизненно важны для снабжения энергией и для продолжения жизни. По сути, цикл Кребса “отвечает” за основной источник энергии во всех живых организмах.
Роль цикла трикарбоновых кислот
Цикл Кребса, также известный как цикл трикарбоновых кислот (TCA), был впервые обнаружен в 1937 году немецким биохимиком Гансом Адольфом Кребсом. Его весьма подробные и обширные исследования в области клеточного метаболизма и других научных исследований принесли ему Нобелевскую премию по физиологии и медицине в 1953 году. Помимо цикла лимонной кислоты, Кребс также идентифицировал цикл мочевины в 1932 году.
Цикл Кребса также известен как цикл лимонной кислоты, потому что лимонная кислота является самым первым продуктом, генерируемым этой последовательностью химических превращений, и она также регенерируется в конце цикла.
Цикл Кребса, вероятно, является наиболее важной частью процесса аэробного дыхания, поскольку он стимулирует образование переносчиков электронов. Они важны, поскольку несут энергию, используемую для создания большого количества АТФ.
Как работает цикл Кребса?
Цикл трикарбоновых кислот происходит только в митохондриальном матриксе, во внутренней части митохондрий.
Цикл Кребса, также называемый циклом лимонной кислоты, является вторым важным этапом окислительного фосфорилирования. После того, как гликолиз расщепляет глюкозу на более мелкие 3-углеродные молекулы, цикл Кребса передает энергию от этих молекул переносчикам электронов, которые будут использоваться в цепи переноса электронов для производства АТФ.
Большинство организмов используют глюкозу в качестве основного источника топлива, но должны расщеплять ее и накапливать энергию в АТФ и других молекулах. Цикл Кребса содержится в митохондриях. Внутри митохондриальной матрицы реакции цикла Кребса добавляют электроны и протоны к ряду электронных носителей, которые затем используются цепочкой переноса электронов для производства АТФ.
Цикл Кребса начинается с продуктов гликолиза, которые представляют собой две трехуглеродные молекулы, известные как пируват. Эта молекула является кислой, поэтому цикл Кребса также называют циклом трикарбоновых кислот. В ходе ряда реакций эти молекулы далее распадаются на диоксид углерода. Энергия от молекул передается другим молекулам, называемым переносчиками электронов. Эти молекулы переносят накопленную энергию в цепь переноса электронов, которая, в свою очередь, создает АТФ.
Первый этап использования глюкозы, гликолиз, производит небольшое количество АТФ, а также молекулы, которые будут обрабатываться с помощью цикла Кребса. Во время гликолиза одна молекула глюкозы расщепляется на две более мелкие трехуглеродные молекулы, пируват. Затем он превращается в ацетил-КоА. Затем ацетил-КоА используется в цикле Кребса для производства нескольких основных продуктов. В свою очередь, эти продукты стимулируют образование АТФ, основного источника энергии клетки.
Подробный расклад цикла Кребса и его взаимодействие с различными процессами организма доступны в первом модуле Школы Anti-Age Expert.
Биологическое окисление. цикл Кребса, этапы, дыхательная цепь
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Биологическое окисление
Указанное выше уравнение для окислительно-восстановительной реакции представляет собой обобщенную форму, так как изображает процесс окисления субстратов как прямое дегидрирование, причем кислород выступает в роли непосредственного акцептора водорода. На самом деле кислород участвует в транспорте электронов иным образом. Существуют промежуточные переносчики при транспорте электронов от исходного донора электронов SH2 к терминальному акцептору – О2. Полный процесс представляет собой цепь последовательных окислительно-восстановительных реакций, в ходе которых происходит взаимодействие между переносчиками. Каждый промежуточный переносчик вначале выступает в роли акцептора электронов и протонов и из окисленного состояния переходит в восстановленную форму. Затем он передает электрон следующему переносчику и снова возвращается в окисленное состояние. На последней стадии переносчик передает электроны кислороду, который затем восстанавливается до воды. Совокупность последовательных окислительно-восстановительных реакций называется цепью переноса (транспорта) электронов, или дыхательной цепью:

Перенос электронов и протонов с участием промежуточных переносчиков. SH2 – исходный донор протонов и электронов; P – промежуточные переносчики; E1, E2, E3, E4 – ферменты окислительно-восстановительных реакций

Последовательность промежуточных переносчиков протонов и электронов в дыхательной цепи
Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы ни был исходный субстрат, электроны и протоны от флавинов переносятся к коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками. NAD – зависимая дегидрогеназа катализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD + является коферментом и выполняет роль акцептора водорода:

Символ 2Н + означает два электрона и два протона, обычно переносимые в виде гидрид иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода». FAD – зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Коферментом является FAD, который является акцептором водорода от субстрата. NADH – дегидрогеназа катализирует окисление NADH и восстановление убихинона (CoQ). Переносчиком водорода является кофермент – FMN (комплекс 1). В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем передается на убихинон. Флавиновые коферменты (FAD и FMN) прочно связаны с ферментом как простетические группы, поэтому ферменты, в состав которых они входят, называются флавопротеины. Флавинмононуклеотид (FMN), или рибофлавин фосфат, неразрывно связан с белковой частью фермента. Строго говоря, FMN не является нуклеотидом, так как флавиновая часть связана с рибитолом, а не с рибозой.
Убихинон (кофермент Q) – производное изопрена:

Название «убихинон» возникло из-за его повсеместной распространенности в природе. Кофермент Q действует как переносчик электронов на цитохромы.
Цитохромы – это гемопротеины – белки, содержащие в качестве прочно связанной простетической группы гем:

Простетическая группа гема в структуре цитохромов
Атом железа в геме может менять валентность, присоединяя или отдавая электроны.В дыхательной цепи цитохромы служат переносчиками электронов и располагаются соответственно величине окислительно-восстановительного потенциала следующим образом: B, С1, С, а, а3. Гемовые группы цитохромов связаны с белковой частью донорно-акцепторными связями между ионом железа и соответствующими аминокислотными остатками:

Связывание гема с белковой частью цитохрома С
В цитохромах С и С1 дополнительные ковалентные связи формируются между тиогруппами цистеина и боковыми винильными группами гема. QН2-дегидрогеназа (комплекс III) представляет собой комплекс цитохромов b и С1. Этот фермент катализирует окисление восстановленного кофермента Q и перенос электронов на цитохром С. Электроны последовательно переносятся атомами железа цитохромов b и С1, а затем поступают на цитохром С. Протоны после окисления QH2 освобождаются в раствор.
Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV). Цитохромоксидаза кроме гема содержит ионы меди, которые способны менять валентность и таким способом участвовать в переносе электронов.
Цитохромоксидаза переносит электроны с цитохрома С на кислород. В переносе электронов участвуют сначала ионы железа цитохромов а и а3, а затем ион меди цитохрома а3. Молекула кислорода связывается с железом в геме цитохрома а3. Следовательно, переход электронов на кислород с иона меди цитохрома а3, происходит на молекуле фермента. Каждый из атомов молекулы кислорода присоединяет по два электрона и протона, образуя при этом молекулу воды.
Белки, содержащие негеминовое железо. Некоторое количество атомов железа в митохондриях связано не в геме цитохромов, а образует комплексы с другими белками. Эти белки называют также железосерными, так как атомы железа связаны с атомами серы цистеиновых остатков. Белки, содержащие негеминовое железо, участвуют в переносе электронов на нескольких стадиях, однако, не совсем ясны их локализация и механизм действия.
Биологическое окисление. Синтез АТФ
Во внутренней мембране митохондрий расположен интегральный белковый комплекс – Н+-зависимая АТФ-синтаза seu Н+-зависимая АТФ-аза (два разных названия связаны с полной обратимостью катализируемой реакции), обладающий значительной молекулярной массой – более, чем 500кДа. Состоит из двух субъединиц: FO и F1.
F1 представляет из себя грибовидный вырост на матриксной поверхности внутренней митохондриальной мембраны, FO же пронизывает эту мембрану насквозь. В толще FO расположен протонный канал, позволяющий протонам возвращаться обратно в матрикс по градиенту их концентраций.
F1 способна связывать АДФ и фосфат на своей поверхности с образованием АТФ – без затраты энергии, но обязательно в комплексе с ферментом. Энергия необходима лишь для освобождения АТФ из этого комплекса. Эта энергия выделяется в результате тока протонов через протонный канал FO.
В дыхательной цепи сопряжение абсолютно: ни одно вещество не может окисляться без восстановления другого вещества.
Но при синтезе АТФ сопряжение одностороннее: окисление может идти без фосфорилирования, а фосфорилирование без окисления никогда не идёт. Это означает, что система МтО может работать без синтеза АТФ, но АТФ не может быть синтезирована, если не работает система МтО.
Биологическое окисление. Сопряжение работы дыхательной цепи с процессом синтеза АТФ
Существование такого сопряжения доказывается тем, что можно ингибировать образование АТФ, не нарушая процесса транспорта электронов. Это достигается добавлением химических веществ, названных разобщителями. После удаления разобщителей синтез АТФ восстанавливается.
Существует несколько гипотез, объясняющих механизм сопряжения. Одной из них является хемоосмотическая теория. Цепь транспорта электронов функционирует как протонная (Н + )помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство. Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи. Перенос протонов приводит к возникновению разности концентрации Н + с двух сторон митохондриальной мембраны: более высокая концентрация будет снаружи и более низкая – внутри. Митохондрия в результате переходит в «энергизованное» состояние, так как возникает градиент концентрации Н + и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности.

С опряжение цепи транспорта электронов и фосфорилирования ADP посредством протонного градиента

С труктура компонентов комплекса I, обеспечивающего функционирование «протонной помпы» при окислении NADH
Биологическое окисление. ОСНОВНЫЕ ПРОЦЕССЫ, ДЛЯ КОТОРЫХ ИСПОЛЬЗУЕТСЯ ЭНЕРГИЯ АТФ:
1. Синтез различных веществ.
2. Активный транспорт (транспорт веществ через мембрану против градиента их концентраций). 30% от общего количества расходуемого АТФ приходится на Na+,К+-АТФазу.
3. Механическое движение (мышечная работа).
Биологическое окисление. Цикл Кребса
Основной функции Цикла Кребса является образование водородных эквивалентов, которые в цепи окислительного фосфорилирования обеспечивают синтез макроэргических соединений.
Кроме того, ЦТК выполняет ведущую роль в процессах глюконеогенеза, переаминирования, дезаминирования АК, липогенеза и синтеза гема. Интегрирует все виды обмена веществ.
Регуляция Цикла Кребса. Осуществляется с участием 4 регуляторных ферментов: цитратсинтазы, изоцитрат ДГ, α-КГ ДГ и СДГ. Цикл Кребса ингибируется в основном НАДН2 и АТФ, которые являются продуктами ЦТК и цепи окислительного фосфорилирования. Активируют цикл Кребса в основном НАД + и АДФ.
Биологическое окисление. Реакции цикла Кребса
1). Цитратсинтаза локализуется в матриксе митохондрий, ее активируют ЩУК, НАД + ; ингибируют АТФ, НАДН2, Сукцинил-КоА, цитрат.

2). Аконитаза локализуется в матриксе митохондрий.

3). Окислительно-восстановительная реакция, самая медленная в ЦТК.

4). Окислительно-восстановительная реакция.

5). Реакция субстратного фосфорилирования

6). Окислительно-восстановительная реакция.
Сукцинатдегидрогеназа, флавопротеин содержащий Fe2S2, связана с внутренней мембраной митохондрии. СДГ ингибирует ЩУК и Сукцинил-КоА,

7). Фумараза локализуется в матриксе митохондрий.

8). Окислительно-восстановительная реакция.
Малат ДГ локализуется в матриксе митохондрий.

Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь.
Биологическое окисление. Энергетический баланс одного оборота ЦТК
В 4 окислительно-восстановительных реакциях ЦТК образуются 3 НАДН2 и 1 ФАДН2, которые направляются далее в дыхательную цепь окислительного фосфорилирования. В процессе окислительного фосфорилирования ДЦ из 1 НАДН2 образуется 3 АТФ, из 1 ФАДН2 – 2 АТФ. Из 1 ГТФ, образующейся в ЦТК за счет субстратного фосфорилирования, синтезируется 1 АТФ. Таким образом, за 1 цикл ЦТК из 3 НАДН2, 1 ФАДН2 и 1 ГТФ получается 12 АТФ.
Биологическое окисление. ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦТК, лимоннокислый цикл, цикл Кребса)
ЦТК, как и реакции митохондриального окисления, протекает в митохондриях. Представляет собой серию реакций, замкнутых в цикл.


Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь от образования цитрата до его превращения в ЩУК.
В реакциях этого цикла участвуют четыре из девяти субстратов МтО.
Происходит серия дегидрогеназных реакций. Из них 3-я, 4-я и 8-я происходят с участием НАД-зависимых дегидрогеназ, и каждая из этих реакций позволяет получить 3 молекулы АТФ. На 6-й стадии происходит ФАД-зависимая дегидрогеназная реакция, которая сопряжена с образованием 2-х молекул АТФ (Р/О = 2).
На 5-й стадии 1 молекула АТФ образуется путем субстратного фосфорилирования.
Итого за 1 оборот ЦТК образуется 12 молекул АТФ.
Смысл ЦТК заключается в том, чтобы остатки уксусной кислоты расщепились с образованием большого количества АТФ. Кроме того, из ацетатных остатков образуется СО2 и Н2О, как конечные продукты обмена веществ.
СО2 образуется в ходе ЦТК дважды:
1. на третьей стадии (окисление изоцитрата)
2. на четвертой стадии (окисление альфа-кетоглутарата).
Если прибавить еще 1 молекулу СО2, которая образуется до начала ЦТК – при превращении ПВК в Ацетил-КоА, то можно говорить о трех молекулах СО2, образующихся при распаде ПВК. Суммарно эти молекулы, образующиеся при распаде ПВК, составляют до 90% углекислоты, которая выводится из организма.
ИТОГОВОЕ УРАВНЕНИЕ ЦТК

БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ЦТК
ГЛАВНАЯ РОЛЬ ЦТК – ОБРАЗОВАНИЕ БОЛЬШОГО КОЛИЧЕСТВА АТФ.
1. ЦТК – главный источник АТФ. Энергию для образования большого количества АТФ дает полный распад Ацетил-КоА до СО2 и Н2О.
2. ЦТК – это универсальный терминальный этап катаболизма веществ всех классов.
3. ЦТК играет важную роль в процессах анаболизма (промежуточные продукты ЦТК):
АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ЦТК
В ЦТК два ключевых фермента:
Оба фермента аллостерически ингибируются избытком АТФ и НАДН2. Изоцитратдегидрогеназа сильно активируется АДФ. Если АДФ нет, то этот фермент неактивен. В условиях энергетического покоя концентрация АТФ увеличивается, и скорость реакций ЦТК мала – синтез АТФ уменьшается.
Изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза, поэтому в условиях энергетического покоя повышается концентрация цитрата, и он выходит в цитоплазму по градиенту концентраций путем облегченной диффузии. В цитоплазме цитрат превращается в Ацетил-КоА, который участвует в синтезе жирных кислот.
Биологическое окисление. ЭНЕРГЕТИЧЕСКИЙ ЗАРЯД КЛЕТКИ (ЭЗК)
Многие процессы метаболизма регулируются соотношением АТФ/АДФ. Это соотношение характеризует энергетический статус (состояние) клетки (Энергетический Заряд Клетки).
Теоретически ЭЗК может меняться от 0 до 1. Обычно в норме величина ЭЗК поддерживается в клетке в пределах 0.85-0.90.
ОБРАТИТЕ ВНИМАНИЕ! Метаболические пути, приводящие к образованию АТФ, ингибируются высоким ЭЗК; метаболические пути, ведущие к использованию АТФ, активируются высоким ЭЗК.
Биологическое окисление. ЧЕЛНОЧНЫЕ МЕХАНИЗМЫ ПЕРЕНОСА ВОДОРОДА
Никотинамидные дегидрогеназы находятся не только в матриксе митохондрий, но и в цитозоле. Митохондриальная мембрана непроницаема для НАД, поэтому НАДН2, который образуется в цитозоле, может передать свой водород в митохондрию только с помощью специальных субстратных ЧЕЛНОЧНЫХ МЕХАНИЗМОВ. В митохондрию из цитозоля передается не сам НАДН2, а только водород, отнятый от него. Переносимый водород включается в молекулу вещества-челнока, способного проникать через митохондриальную мембрану. В митохондрии вещество-челнок отдает водород на митохондриальный НАД или ФАД и возвращается обратно в цитозоль.
 В клетках организма человека существуют 2 типа челночных механизмов.
В клетках организма человека существуют 2 типа челночных механизмов.
1. МАЛАТ-АСПАРТАТНЫЙ челнок (наиболее универсален для клеток организма). С высокой скоростью работает в миокарде, почечной ткани, печени.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный НАД(!), поэтому в митохондриях образуется 3 молекулы АТФ и не происходит потери энергии при переносе водорода. Для ткани печени малат-аспартатная система особенно важна, так как из митохондрии выводится Ацетил-КоА (в виде цитрата), а водород попадает в митохондрию (в составе малата).
 Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
ЩУК может вернуться в цитоплазму и другим способом: она может вступить в реакцию трансаминирования с глутаминовой кислотой (СМОТРИТЕ РИСУНОК)
2. ГЛИЦЕРОФОСФАТНЫЙ челнок (встречается реже).
 В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х – происходит потеря энергии при переносе водорода.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х – происходит потеря энергии при переносе водорода.
В клетке существует не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному. Происходит и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
Биологическое окисление. Пути использования кислорода
Биологическое окисление. Историческая справка
Изучение окисления в организме было начато в 18 в. А. Лавуазье; в дальнейшем значительный вклад в исследование БО (его локализация в живых клетках, связь с др. процессами обмена веществ, механизмы ферментативных окислительно-восстановительных реакций, аккумуляция и превращение энергии и др.) внесли О. Варбург, Г. Виланд (Германия), Д. Кейлин, Х. Кребс, П. Митчелл (Великобритания), Д. Грин, А. Ленинджер, Б. Чанс, Э. Рэкер (США), а в СССР – А. Н. Бах, В. И. Палладин, В. А. Энгельгардт, С. Е. Северин, В. А. Белицер, В. П. Скулачев и др.
В 1777г французский химик Антуан Лоран Лавуазье (1743-1794), впервые правильно истолковал явление горения как процесс соединения веществ с О2. Так как горение и дыхание сопровождаются потреблением О2 и выделение СО2, он предположил что, в их основе лежит один процесс. Однако у дыхания были существенные отличия от горения: оно шло при низкой температуре, без пламени и в присутствии воды.
В конце XIX века русские исследователи А.Н. Бах и В.И. Палладин, работая независимо друг от друга, предложили 2 основные теории для объяснения процессов, протекающих в ходе биологического окисления.
В 1897г Алексей Николаевич Бах (1857-1946) сформулировал «перекисную теорию окисления», согласно которой молекула О2 сначала активируется. Активация О2 происходит в результате разрыва одной его связи (-О-О-) и присоединения к органическим веществам – оксидазам. Активированный О2 при взаимодействии с окисляемым веществом образует перекись.

Теория «перекисного окисления» Баха нашла свое подтверждение, однако главный механизм БО оказался иным.
Важнейшая заслуга в развитии учения о БО принадлежит русскому ученому ботанику и биохимику В.И. Палладину (1859-1922), который создал теорию «активации водорода». Он предположил, что окисление субстратов может происходить в 2 фазы:
1). Анаэробная фаза. В этой фазе особые вещества хромогены (R) отщепляют Н от субстратов и восстанавливаются (RH2).
2). Аэробная фаза. Восстановленные хромогены RH2 передают Н на О2.

В последствии теория В.И. Паладина подтвердилась для процессов митоходриального окисления, а хромогены, принимающие участие в отнятии водорода от субстратов, в настоящее время называются ферментами дегидрогеназами.
Биологическое окисление. Современные представления
Согласно современной теории БО:
Окислительно-восстановительные реакции (ОВР) – реакции, в которых меняется степень окисления субстрата за счет присоединения/отщепления: 1) 1 е – ; 2) 2е – и 2Н + ; 3) атомов кислорода.
Биологическое окисление (БО) совокупность окислительно-восстановительных реакций, которые протекают во всех живых клетках.
Субстрат БО – вещество, способное отдавать электрон. (Любые вещества, способные вступать в реакции окисления).
Тканевое дыхание – окисление органических веществ в клетках, сопровождающееся потреблением О2 и выделением воды.
Субстрат тканевого дыхания – это вещество, которое отдает электрон непосредственно в цепь окислительного фосфорилирования. (Например, НАДН2, ФАДН2).
Дыхательная цепь – цепь переноса электронов. В переносе электронов от субстратов БО к О2 принимают участие: 1) НАД и НАДФ зависимые ДГ; 2) ФАД и ФМН зависимые ДГ; 3) цитохромы; 4) коэнзим Q; 5) белки, содержащие негеминовое железо.
Свободная энергия. Каждое органическое вещество обладает определенным запасом внутренней энергии (Е). Часть этой внутренней энергии может быть использована для совершения полезной работы, такую энергию называют свободной (G). Направление химической реакции определяется значением ∆G. У катаболических реакций ∆G отрицательно, эти реакции протекают самопроизвольно (экзергонические реакции). У анаболических реакций ∆G положительно, они протекают только при поступлении свободной энергии извне (эндергонические реакции).
Редокс-потенциалы. В каждой окислительно-восстановительной системе участвует окисленная и восстановленная формы одного соединения, которые образуют сопряженную окислительно-восстановительную или редокс-пару. Разные редокс-пары обладают различным сродством к электрону. Мерой сродства редокс-пары к электрону служит окислительно-восстановительный потенциал, или редокс-потенциал (Ео‘), величина которого прямо пропорциональна изменению свободной энергии ∆G. Величину Ео‘ выражают в вольтах; чем она отрицательнее, тем меньше сродство вещества к электронам и наоборот.
Биологическое окисление. Этапы унифицирования энергии пищевых веществ и образования субстратов тканевого дыхания
Образование субстратов тканевого дыхания осуществляется в несколько этапов:
1. При переваривании в ЖКТ происходит гидролиз полимеров (белков, полисахаридов) и ТГ до мономеров, которые потом всасывающихся в кровь и включающихся в промежуточный обмен.
2. В ходе катаболизма мономеры (моносахара, жирные кислоты и аминокислоты) превращаются в универсальное вещество – Ацетил-КоА (исключение некоторые АК).
3. Ацетил-КоА поступает в ЦТК, где из него в последовательных реакциях образуются субстраты тканевого дыхания: изоцитрат, α-КГ, сукцинат и малат.
4. Окисление субстратов тканевого дыхания сопровождается восстановлением коферментов НАДН2 и ФАДН2, которые затем отдают протоны в редокс-цепь окислительного фосфорилирования.
Биологическое окисление. Пути использования кислорода в клетке
В настоящее время выделено 4 основные пути использования кислорода в организме:
1. Оксидазный путь – окислительное фосфорилирование. Протекает в митохондриях, является основным источником АТФ в аэробных тканях. Потребляет 90% кислорода.
2. Монооксигеназный путь. Обеспечивает включение 1 атома кислорода в молекулу субстрата. Используется для синтеза новых веществ (стероидные гормоны), обезвреживания ксенобиотиков и токсических продуктов обмена в митохондриях и ЭПР.
3. Диоксигеназный путь. Обеспечивает включение молекулы кислорода в молекулу субстрата. Используется для деградации АК и синтеза новых веществ.
4. Пероксидазный и радикальный пути. Кислород участвует в образовании перекисей и активных радикалов, которые необходимы в пероксисомах для внутриклеточного пищеварения, разрушения макрофагами бактерий, вирусов, регуляции метаболизма и т.д. Перекиси и активные кислородные радикалы оказывают также повреждающее воздействие на структуры клеток и тканей, активируя ПОЛ. Разрушение перекисей и инактивация свободных радикалов осуществляется с помощью ферментативной и неферментативной антиокидантной системы.
Биологическое окисление. Типы оксидоредуктаз
В соответствие с путями потребления кислорода и катализируемыми реакциями, все оксидоредуктазы разделены на 5 групп:
1). Оксидазы удаляют Н из субстрата (S), используя в качестве акцептора Н только О2, продуктом реакции является вода. Все оксидазы содержат медь. Например, цитохромоксидаза, последний фермент дыхательной цепи.
2). Аэробные дегидрогеназы, или ФАД зависимые оксидазы, относятся к металлофлавопротеинам (ФАД, ФМН, Fe, Cu, Mo), находятся в пероксисомах и наружной мембране митоходрий. Они отнимают Н от субстрата, и передают его на О2 с образованием перекиси. Дегидрогеназа L-аминокислот (оксидаза L-аминокислот) в почках осуществляет окислительное дезаминирование. Ксантиндегидрогеназа (ксантиноксидаза) превращает пурины в мочевую кислоту. Моноаминоксидазы (МАО) окисляют гормон адреналин и некоторые биогенные амины. Диаминоксидазы (ДАО) окисляют гистамин и другие диамины и полиамины. Образующаяся перекись бактерицидное действие.
3). Анаэробные дегидрогеназы многочисленная группа, содержат в качестве коферментов НАДН2, НАДФН2, ФАД, ФМН, цитохромы. Они удаляют Н из субстрата, не используя в качестве акцептора протона О2. Анаэробные дегидрогеназы выполняют две главные функции: перенос Н с одного субстрата на другой в сопряженной ОВР и транспорт электронов в дыхательной цепи. Примеры ферментов: изоцитрат ДГ (НАДН2), сукцинат ДГ (ФАДН2), цитохромы в, с1, с, а и а3.
4). Оксигеназы. Включают кислород в молекулу субстрата. Работают в составе мультиферментного комплекса, содержащего ФАД зависимую ДГ, Fe2S2-белок, цитохромы Р450 или В5. Этот комплекс встроен в мембрану ЭПР или внутреннюю мембрану митохондрий.
а) Монооксигеназы. Обеспечивают включение 1 атома кислорода в молекулу субстрата. В качестве восстановителей (R) используется НАДФН2, НАДН2 и аскорбат. Участвуют в синтезе стероидных гормонов, обезвреживании ксенобиотиков. Фен-4-монооксигеназа окисляет фен в тир. Пролингидроксилаза (аскорбат) гидроксилирует пролин в проколлагене.
б) Диоксигеназы. Обеспечивают включение молекулы кислорода в молекулу субстрата, окисляют циклические соединения с разрывом цикла. Например, триптофаноксигеназа, превращает триптофан в фенилкенуренин.
5). Гидроксипероксидазы разрушают перекиси водорода и органические перекиси. Ферменты пероксидаза и каталаза. Пероксидазы, содержат протогем, восстанавливают перекись до воды за счет хинонов, аскорбата или цитохрома с. Глутатионперосидаза, содержит селен, восстанавливает перекись до воды за счет глутатиона.
Биологическое окисление. Варианты дыхательной цепи
1. ПОЛНАЯ ДЫХАТЕЛЬНАЯ ЦЕПЬ
В этой цепи окисляется небольшое количество субстратов, из которых главными являются четыре. Коэффициент Р/О=3.
| СУБСТРАТЫ | НИКОТИНАМИДНЫЕ ФЕРМЕНТЫ, ИХ ОКИСЛЯЮЩИЕ |
| изолимонная кислота (изоцитрат) | Изоцитратдегидрогеназа |
| Яблочная кислота (малат) | Малатдегидрогеназа |
| Глутаминовая кислота (глутамат) | Глутаматдегидрогеназа |
| бета-гидроксиацил-КоА | бета-гидроксиацил-КоА-дегидрогеназа |
Все ферменты полной цепи являются НАД-зависимыми дегидрогеназами.
2. СОКРАЩЕННАЯ (УКОРОЧЕННАЯ) ДЫХАТЕЛЬНАЯ ЦЕПЬ
Известны два субстрата. Главным из них является сукцинат (янтарная кислота).

Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи – исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования – коэффициент Р/О=2.
Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты):

3. МАКСИМАЛЬНО СОКРАЩЕННАЯ (МАКСИМАЛЬНО УКОРОЧЕННАЯ) ДЫХАТЕЛЬНАЯ ЦЕПЬ.
Она представлена только цитохромной частью. Эксперименты показали, что здесь может быть окислен только один субстрат – аскорбиновая кислота, с участием фермента, восстановленные эквиваленты включаются в цепь на уровне цитохрома С цитохромоксидазы (цитохром аа3), но в реальных условиях такого окисления практически не происходит. Образуется вода и 1 молекула АТФ. Коэффициент Р/О=1.
Возможность такого окисления доказана в эксперименте in vitro. А в живой клетке аскорбиновая кислота обычно используется как донор водорода в системе окисления оксигеназного типа (реакции, катализируемые монооксигеназами: смотрите лекцию «Внемитохондриальное окисление»). Такие реакции с участием витамина «С» особенно важны для формирования белка коллагена, в котором за счет монооксигеназной реакции образуется гидроксипролин.
Биологическое окисление. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРОВИНОГРАДНОЙ И a-КЕТОГЛУТАРОВОЙ КИСЛОТ В МИТОХОНДРИЯХ
Этот вариант дыхательной цепи удлинен по сравнению с полной цепью за счет того, что первое звено катализируется не никотинамидным ферментом, а мультиферментным комплексом. Это единая надмолекулярная структура. В состав этого комплекса входят 3 фермента и 5 коферментов. Такой комплекс называется мультиферментным комплексом окислительного декарбоксилирования a-кетокислот, и он окисляет два субстрата:
1. Пировиноградную кислоту (пируват, ПВК)
Окисляется с помощью ферментов пируватдегидрогеназного комплекса;
2. a-кетоглутаровую кислоту (a-КГ)
Окисляется с помощью ферментов a-кетоглутаратдегидрогеназного комплекса.
Оба комплекса ферментов работают одинаково. Они катализируют реакции окислительного декарбоксилирования соответствующей a-кетокислоты.
Биологическое окисление. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
1-ю реакцию катализирует фермент ПИРУВАТДЕКАРБОКСИЛАЗА (Е1).
Простетической группой пируватдекарбоксилазы является тиаминдифосфат (ТПФ, тиаминпирофосфат, ТДФ) – это активная форма витамина В1. Активная часть ТПФ – тиазоловое кольцо и атом водорода в нем. Для краткости записывают: НС-ТПФ.

Пируватдекарбоксилаза отщепляет CO2, а оставшаяся оксиэтильная группа присоединяется к ТПФ.


2-ю и 3-ю реакцию процесса катализирует фермент АЦИЛТРАНСФЕРАЗА (Е2). Простетическая группа ацетилтрансферазы – амид липоевой кислоты. Катализирует перенос оксиэтильного остатка на свой собственный кофермент (строение ЛК знать описательно по учебнику). В ее составе есть дисульфидная связь.

На этом этапе уже произошло окисление оксиэтильного остатка до остатка уксусной кислоты, одновременно с этим началось восстановление липоевой кислоты.
3 этап – продолжает работать фермент ацилтрансфераза.
Выяснено, что этот фермент переносит не только остатки уксусной кислоты, но и остатки других кислот (нециклических). Поэтому его еще называют коферментом ацилирования.

Второй фермент данного комплекса является не только ацетилтрансферазой, но и окислительным ферментом одновременно (дегидрогенизирующая ацетилтрансфераза).
Энергия этого окисления аккумулируется в виде макроэргической связи активной формы уксусной кислоты (ацетил-КоА).
4 этап катализируется ферментом ДИГИДРОЛИПОИЛДЕГИДРОГЕНАЗОЙ.
Кофермент этого этапа – особый ФАД с низким окислительно-восстановительным потенциалом (обозначается как ФАД’). Переносит протоны и электроны с дигидролипоата на НАД – последний кофермент в описанном комплексе.

В итоге можем записать суммарное уравнение:

Аналогично действует ферментный комплекс для окислительного декарбоксилирования a-кетоглутарата. Но названия двух из трех ферментов другие:
1-й фермент – декарбоксилаза a-кетоглутаровой кислоты
2-й фермент – сукцинилтрансфераза (переносит остаток янтарной кислоты)
Суммарное уравнение для этого комплекса:

В результате действия этих двух комплексов, каждый из двух приведенных субстратов (ПВК и a-КГ) – теряют СО2, два протона и два электрона, которые передаются на кислород с образованием Н2О и параллельно происходит фосфорилирование трех молекул АТФ. Сами субстраты превращаются в итоге в активную форму соответствующей кислоты.
Макроэргическая связь в молекулах этих активных форм может легко трансформироваться в макроэргическую связь молекулы АТФ. Например, для молекулы сукцинил-КоА:

Такой способ образования АТФ, когда нет сопряжения с процессами окисления, называется СУБСТРАТНЫМ ФОСФОРИЛИРОВАНИЕМ (при этом используется макроэргическая связь в молекуле какого-нибудь субстрата). В организмах высших животных и человека этот путь получения АТФ не является главным. Благодаря такому механизму окислительное декарбоксилирование a-кетокислот может считаться энергетически более выгодной, потому что образуется вещество с макроэргической связью – ацетил-КоА. Некоторые синтетазы используют макроэргические связи из Ацетил-КоА или Сукцинил-КоА для реакций биосинтеза.
Например, при синтезе гема гемоглобина используется часть молекул сукцинил-КоА:

Большая часть будет все же использоваться для синтеза ГТФ.
Другой продукт окислительного декарбоксилирования (Ацетил-КоА) может расходоваться в реакциях синтеза жирных кислот, холестерина, стероидов. А самая большая часть этого вещества используется при синтезе лимонной кислоты в реакциях Цикла Трикарбоновых Кислот.
Биологическое окисление. ЭНЕРГЕТИЧЕСКИЙ ИТОГ УДЛИНЕННОЙ ЦЕПИ.
Образуются 3 молекулы АТФ путем окислительного фосфорилирования. Продукты окислительного декарбоксилирования (ацетил-КоА и сукцинил-КоА) содержат макроэргическую связь. Энергия макроэргической связи сукцинил-КоА используется двумя способами.
Энергия макроэргической связи Ацетил-КоА используется только в реакциях синтеза:
Среди субстратов МтО почти нет веществ, которые организм получает с пищей – только глутаминовая кислота.
Субстраты МтО – в основном органические кислоты, которые образуются в организме в процессе катаболизма. В процессе катаболизма из большого количества разнообразных пищевых веществ образуется всего 2 вида общих метаболитов: Ацетил-КоА и ПВК.
Биологическое окисление. Пути использования кислорода
Биологическое окисление. ОКСИГЕНАЗНЫЙ ПУТЬ
Оксигеназы это ферменты, которые катализируют включение кислорода в молекулу субстрата. Оксигеназы работают в составе мультиферментных комплексов, встроенных в мембрану и участвуют в синтезе и деградации многих типов метаболитов. По способу включения кислорода их делят на: монооксигеназы и диоксигеназы.
Монооксигеназные реакции
Монооксигеназы это ферменты, которые включают в субстрат только один атом молекулы кислорода. Другой атом кислорода восстанавливается до воды с участием электронов и протонов НАДФН2, НАДН2, реже витамин С:
Монооксигеназные реакции протекают на цитоплазматической поверхности гладкого ЭПР, их называют микросомальным окислением, и на внутренней поверхности внутренней мембраны митохондрии.
1. Микросомальные монооксигеназные системы
Катализируют низкоспецифичные реакции. Эти монооксигеназы функционируют в комплексе с различными ЦПЭ:
А. Цепь НАДФН2-Р450 редуктаза – Цитохром Р450
Донорами протонов и электронов для этой цепи являются НАДФН2.
Субстратами являются гидрофобные вещества экзогенного (лекарства, ксенобиотики) и эндогенного (стероиды, жирные кислоты и т.д.) происхождения.
Регуляция активности осуществляется индукцией синтеза ферментов. Открыто более 250 веществ-индукторов (барбитураты, спирты, кетоны, стероиды, ароматические углеводороды).

.НАДФН2-Р450 редуктаза. Цитозольный домен содержит 2 кофермента ФАД и ФМН, гидрофобный домен фиксирует фермент в мембране. НАДФН2-Р450 редуктаза переносит электроны с НАДФН2 на цитохром Р450.
Цитохром Р450 – интегральный гемопротеин, содержит простетическую группу гем, имеет участки связывания для О2 и субстрата. Открыто 150 генов, кодирующих различные изоформы цитохрома Р450. Каждая из изоформ Р450 имеет много субстратов и отличается от других изоформ Р450 только белковой частью.
Б. Цепь НАДН2-цитохром b5 редуктаза – Цитохром b5 – стеароил-КоА-десатураза
Донорами протонов и электронов для этой цепи являются НАДН2.

НАДН2-цитохром b5 редуктаза – двухдоменный белок, цитозольный домен содержит ФАД, гидрофобный домен фиксирует фермент в мембране. НАДН2-b5 редуктаза переносит электроны с НАДН2 на цитохром b5.
Цитохром b5. Цитозольный домен содержит гем, гидрофобный домен фиксирует фермент в мембране. Цитохром b5 может передавать свои электроны на различные ферменты (цитохром Р450, Стеароил-КоА-десатуразу и т.д.), образуя различные ЦПЭ, при этом он участвует в десатурации и элонгации жирных кислот, в синтезе холестерина, плазминогенов и церамида.
Стеароил-КоА-десатураза – интегральный фермент, содержит негеминовое железо. Катализирует образование 1 двойной связи между 9 и 10 атомами углерода в жирных кислотах. Стеароил-КоА-десатураза переносит электроны с цитохрома b5 на 1 атом кислород, при участии протонов этот кислород образует воду. Второй атом кислорода включается стеариновую кислоту с образованием её оксиацила, который дегидрируется до олеиновой кислоты.
Биологическое окисление. Митохондриальные монооксигеназные системы
Митохондриальные монооксигеназные системы локализованы на внутренней поверхности внутренней мембране митохондрий и катализируют высокоспецифичные реакции.
Компонентами этих систем могут быть: НАДН2-зависимые ФАД-содержащее редуктазы, Fe2S2-белки (адренодоксин), цитохромы Р450, b5, элонгазы и т.д.

Митохондриальные монооксигеназные системы:
· в коре надпочечников, семенниках, яичниках и плаценте участвуют в синтезе стероидных гормонов из холестерина (гидроксилирование по С22 и С20 при отщеплении боковой цепи и по положениям 11β и 18);
· в почках участвуют в синтезе 1,25-диоксихолекальциферола (Витамин Д3) из 25-гидроксихолекальциферола;
· в печени участвуют в синтезе желчных кислот из холестерина;
· в соединительной ткани участвуют в гидроксилировании пролина в оксипролин в молекуле коллагена (пролингидроксилазы). Донором протонов и электронов является витамин С. С оксипролином зрелый коллаген приобретает механическую прочность.
Биологическое окисление. Диоксигеназные реакции
Диоксигеназы это ферменты, которые включают в субстрат оба атома молекулы кислорода:
Таким путем окисляются циклические трудноокисляемые структуры, реакции идут с разрывом цикла. Диоксигеназные реакции протекают на цитоплазматической поверхности гладкого ЭПР.

L-триптофандиоксигеназа печени, содержит гем, участвует в катаболизме триптофана:

Биологическое окисление. ПЕРОКСИДАЗНЫЙ И РАДИКАЛЬНЫЙ ПУТИ ИСПОЛЬЗОВАНИЯ КИСЛОРОДА
Кислород – потенциально опасное вещество. Молекулярный О2 является бирадикалом (˙О::О˙), но он стабилен так как, 2 неспаренных е – внешней орбиты имеют параллельные спины, (подавляют радикальную активность друг друга).
Кислород в составе молекулы Н2О стабилен, потому что его внешняя электронная орбита укомплектована е – (Н:О:Н). Химические соединения, в составе которых кислород имеет промежуточную степень окисления, имеют высокую реакционную способность и называются активными формами кислорода (АФК). К активным формам кислорода относятся свободные радикалы кислорода и перекиси. Свободный радикал – свободный атом или частица с неспаренным электроном.
Образование активных форм кислорода
АФК во многих клетках образуются в основном в ферментативных и неферментативных реакциях в результате последовательного присоединения е – к кислороду:
1) О2 + 1е – → О ∙ 2 супероксидный анион-радикал (˙О::О:).
2) О ∙ 2 +1е – → О 2- 2 пероксидный анион (:О::О:), он быстро протонируется с образованием перекиси водорода О 2- 2 + 2Н + → Н2О2 (Н:О::О:Н)
3) Н2О2 + 1е – → НО ∙ + ОН – гидроксильный радикал, ОН – протонируется с образованием воды ОН – + Н + → Н2О
4) ОН ∙ + 1е – → Н2О (Н:О:Н)
В организме донорами электронов являются:
1). ЦПЭ. Утечка электронов из ЦПЭ на кислород является основным путем образования активных форм кислорода в большинстве клеток:
b) в монооксигеназных реакциях электрон с цитохрома Р450 переходит на кислород с образованием супероксидного анион-радикала, который иногда теряется с активного центра.
c) Аэробные дегидрогназы (ФАД-зависимые оксидазы) переносят электроны и протоны с субстрата на кислород с образованием перекиси водорода. Примеры таких оксидаз — оксидазы аминокислот, супероксид дисмутаза, оксидазы, локализованные в пероксисомах.
2). Металлы переменной валентности. Наличие в клетках Fe 2+ или ионов других переходных металлов катализирует образования гидроксильных радикалов и других активных форм кислорода. Например, в эритроцитах окисление иона железа гемоглобина способствует образованию супероксидного анион-радикала.
Hb(Fe 2+ ) + O2 → MetHb(Fe 3+ ) + О ∙ 2
H2O2 + Fe 2+ → Fe 3+ + HO – + HO· (реакция Фентона)
HOCl + Fe 2+ → Fe 3+ + Cl – + HO· (реакция Осипова)
3). Радикалы. Активные формы кислорода, обмениваясь электроном, легко переходят друг в друга: О ∙ 2 + Н2О2 → О2 + НО ∙ + ОН –
Биологическое окисление. Ферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать ЦПЭ. Утечка е – из ЦПЭ на кислород является основным путем образования АФК в большинстве клеток:
Неферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать:
1). Металлы переменной валентности. Наличие в клетках Fe 2+ или ионов других переходных металлов катализирует образования АФК. Например, в эритроцитах окисление иона железа гемоглобина способствует образованию супероксидного анион-радикала.
Hb(Fe 2+ ) + O2 → MetHb(Fe 3+ ) + О ∙ 2
H2O2 + Fe 2+ → Fe 3+ + HO – + HO· (реакция Фентона)
HOCl + Fe 2+ → Fe 3+ + Cl – + HO· (реакция Осипова)
2). Радикалы. АФК, обмениваясь электроном, легко переходят друг в друга: О ∙ 2 + Н2О2 → О2 + НО ∙ + ОН –
Биологическое окисление. Свойства активных форм кислорода
Кислородные радикалы, обладая высокой активностью, разрушают органические молекулы в реакциях свободно-радикального окисления (СРО). Большая часть этих реакций протекает с полиненасыщенными жирными кислотами липидов, и называется перекисным окислением липидов (ПОЛ). Реакции ПОЛ являются цепными.
Наиболее химически активным соединением является гидроксильный радикал – сильнейший окислитель. Время его жизни очень короткое (1 миллиардная доля секунды), но за это время он мгновенно вступает в цепные окислительные реакции в месте своего образования.
Супероксидный анион-радикал и перекись водорода более стабильные вещества, могут диффундировать от места образования, проникать через мембраны клеток. Однако, перекись водорода способствует образованию гидроксильного радикала по следующей реакции:
Fe 2+ + Н2О2 → Fe 3+ + НО ∙ + ОН –
Биологическое окисление. Использование активных форм кислорода в организме
1. Иммунная система. АФК используются фагоцитарными клетками – тканевыми макрофагами, моноцитами и гранулоцитами крови для разрушения бактерий, вирусов и онкоклеток.
В условиях патологии могут произойти нарушения либо системы защитных ферментов (в частности, снижение активности СОД), либо ферментных систем, связывающих ионы железа в плазме крови (церулоплазмин и трансферрин) и в клетках (ферритин). В этом случае супероксидные радикалы и перекись водорода вступают в альтернативные реакции: Образование двухвалентного железа из трехвалентного: Fe 3+ + O •- → Fe 2+ + O2
Реакция перекиси водорода и гипохлорита с ионами двухвалентного железа:
При дефиците в клетках СОД, ферритина, а в плазме церулоплазмина, трансферрина активируются альтернативные реакции:
Фактор некроза опухоли
2. Поддержание гомеостаза.
Эйказаноиды – медиаторы воспаления
3. Внутриклеточное пищеварение. В пероксисомах образуются АФК. Когда пероксисомы сливаются с фагосомами, АФК обеспечивают внутриклеточное пищеварение.
Биологическое окисление. Повреждающее действие активных форм кислорода в организме
Радикалы гидроксила химически исключительно активны и вызывают повреждение белков, нуклеиновых кислот и липидов биологических мембран. В белках происходит неферментативное окисление аминокислотных остатков гистидина, цистеина, триптофана. Таким образом, инактивируют многие ферменты. Особенно тяжелые последствия имеют повреждение ДНК и липидов. Радикалы НО ∙ вызывают разрыв нитей ДНК, обладая, в зависимости от ситуации, мутагенным, канцерогенным или цитостатическим действием. Радикалы гидроксила инициируют цепную реакцию ПОЛ, при этом нарушаются физико-химические свойства мембран – проницаемость, рецепторная функция и работа мембранных белков. Повреждение барьера приводит к нарушению регуляции внутриклеточных процессов и тяжелым расстройствам клеточных функций. Свободно-радикальные реакции часто вызывают гибель клеток и целом ускоряют процесс старения организма.
Биологическое окисление. Антиоксидантные клетки
Антиоксидантная система
В нормальных условиях процесс СРО находится под строгим контролем ферментативных и неферментативных систем клетки, от чего скорость его невелика. Химические соединения и физические воздействия, влияющие на скорость СРО, делят на прооксиданты и антиоксиданты.
Прооксиданты усиливают процессы СРО. Это высокие концентрации кислорода (например, при длительной гипербарической оксигенации больного), ферментные системы, генерирующие супероксидные радикалы (например, ксантиноксидаза, ферменты плазматической мембраны фагоцитов и др.), ионы двухвалентного железа.
Антиоксиданты тормозят СРО. Антиоксиданты, находящиеся в организме, образуют его ферментативную и неферментативную антиоксидантную систему.
1. Ферментативная антиоксидантная система
К ферментам, защищающим клетки от действия активных форм кислорода, относят супе-роксиддисмутазу, каталазу и глутатионпероксидазу. Наиболее активны эти ферменты в печени, надпочечниках и почках, где содержание митохондрий, цитохрома Р450 и пероксисом особенно велико.
Супероксиддисмутаза (СОД) превращает супероксидные анионы в перекись водорода:
Изоферменты СОД находятся и в цитозоле (Cu 2+ и Zn 2+ ) и в митохондриях (Mn 2+ ) и являются как бы первой линией защиты, потому что супероксидный анион образуется обычно первым из активных форм кислорода. СОД — индуцируемый фермент, т.е. синтез его увеличивается, если в клетках активируется СРО.
Каталаза – геминовый фермент, катализирует реакцию разрушения перекиси водорода. При этом образуется вода и молекулярный кислород:
Каталаза находится в основном в пероксисомах, где образуется наибольшее количество перекиси водорода, а также в лейкоцитах, где она защищает клетки от последствий «респираторного взрыва» и в эритроцитах, где она защищает гем гемоглобина от окисления.
Глутатионпероксидаза — обеспечивает разрушение перекиси водорода и гидропероксидов липидов при окислении глутатиона (у-глутамилцистеинилглицин): Н2О2 + 2 GSH → 2 Н2О + G-S-S-G. Глутатионпероксидаза в качестве кофермента содержит селен.
Глутатионредуктаза восстанавливает окисленный глутатион с участием НАДФН2:
Недостаток глутатиона в клетках, например эритроцитах, который может быть обусловлен действием токсических веществ, например ионами тяжелых металлов или наследственным недостатком глутатионредуктазы приводит к активации перекисного окисления; это, в частности, наблюдается при некоторых видах гемолитических анемий.
Фосфолипаза в мембране отщепляет от фосфолипидов окисленные жирные кислоты, содержащие гидроперекисную группу (LOOH), тем самым разрушаются гидроперекиси липидов, предотвращается разветвление цепей окисления липидов в мембранах.
2. Неферментативная антиоксидантная система
“Липидные антиоксиданты” – производные фенола, способны инактивировать свободные радикалы в гидрофобном слое мембран и предотвращать развитие ПОЛ. К ним относится α-токоферол (витамин Е), убихинон (коэнзим Q), тироксин и синтетические соединения, например ионол (бутилированный гидрокситолуол).

Регенерацию α-токоферола осуществляет витамин С.
Свободный радикал витамина Е стабилен и не поддерживает ПОЛ, он взаимодействует с радикалами липидных перекисей, восстанавливает их, а сам превращается в стабильную окисленную форму — токоферолхинон.

Витамин С ингибирует СРО с помощью двух различных механизмов:
1). восстанавливает в мембранах токоферолхинон до витамина Е:
НО-аскорбат-ОН + α-ТФ-О· → α-ТФ-ОН + НО-аскорбат-О· (семидегидроаскорбиновая к-та)
НО-аскорбат-О· + α-ТФ-О· → α-ТФ-ОН + О=аскорбат=О (дегидроаскорбиновая к-та)
Регенерация аскорбиновой кислоты идет с участием ферментативных систем:
а). В микросомах, с участием комплекса НАДН2-редуктаза-цитохром b5:
2НО-аскорбат-О· + НАДН2 → 2НО-аскорбат-ОН + НАД +
б). В митохондриях, с участием НАДН2-семидегидроаскорбатредуктазы:
2НО-аскорбат-О· + НАДН2 → 2НО-аскорбат-ОН + НАД +
в). В цитозоле, с участием НАДФН2-дегидроаскорбатредуктазы:
О=аскорбат=О + 2НАДФН2 → НО-аскорбат-ОН + 2НАДФ +
г). В цитозоле, с участием GSH-дегидроаскорбатредуктазы:
О=аскорбат=О + 2GSH → НО-аскорбат-ОН + GS-SG
2). взаимодействует с активными формами кислорода — О ∙ 2, Н2О2, НО ∙ и инактивирует их.
β-Каротин, предшественник витамина А, также обладает антиоксидантным действием и ин-гибирует ПОЛ.
Соединения, связывающие железо. Большинство из них, включая такие природные соединения как дипептид карнозин, не просто связывают железо, но, самое главное, не дают ему возможности приникнуть в липидную фазу мембран, поскольку образующиеся комплексы, в силу своей полярности, не проникают в гидрофобную зону.
Для детоксикации двухвалентного железа в организме существует, по-видимому, целая система окисления и связывания ионов железа. В плазме крови эта система представлена ферментом церрулоплазмином (феррооксидазой), который окисляет Fe 2+ до Fe 3+ кислородом без образования свободных радикалов, и белком трансферрином, который связывает и переносит в кровяном русле ионы трехвалентного железа, которые затем захватывается клетками. В клетках железо может восстанавливаться аскорбиновой кислотой и другими восстановителями, но затем окисляется и депонируется в окисленной форме внутри ферментного белкового комплекса ферритина.